









Saving the best for last..... Mortimer
spirits in the material world - the police - ghost in the machine album
There is no political solution
To our troubled evolution
Have no faith in constitution
There is no bloody revolution
We are spirits in the material world
Our so-called leaders speak
With words they try to jail you
They subjugate the meek
But it's the rhetoric of failure
We are spirits in the material world
Where does the answer lie?
Living from day to day
If it's something we can't buy
There must be another way
by I.J. Good who may have known of Yakir's 1964 paper. Of course my first introduction to
back-from-the-future physics was in 1953 as a child in a series of mysterious telephone calls.
President Obama recently gave Yakir a medal for it at The White House - well deserved.
This is an important idea that will change theology as well as physics.
http://www.stardrive.org/
Mysterious phone calls? we'll get back to that but first let me introduce you to Mortimer.
Mortimer is a kind of quite fellow as well. As guides go his function as my "guide" was "financial accountant".
Now this threw me for awhile. I just could not see where money/finance had a place in inter-dimensional guides. Made absolutely no sense to me. Now I thought for a time perhaps this guide would help me with my money and guide me to have abundance. For in fact I was making a pretty good income as a Finance Director. But what in heavens name did that have to do with any of this stuff? What was it that had "value" in a financial sense.
So I looked at the wealthiest and realized most of the truly wealthy retainded one thing they "prized" of value. Money was certainly a way of getting places in a material world. But what was of MORE importance to these "elite" types....you know...the ones who ruled Egypt...the ones who are said to rule kingdoms and basically the world? Seems to me "bloodline" took presidency in monarchies and henceforth were handed down from one member of that line to another eh? Apparently money was the "result" of the thing that had REAL value.... and that was a genetic line. Just as the Merovingian's about bloodline. Or the Queen of England. Ask the elusive "33" what they value and try to get an answer why bloodline/genetics seem so important?
So meet Mortimer my financial accountant....and find out how he balances his books in things of financial "wealth".
"Mortimer" Maps - Edition 12

Chromosome I
Chromosome II
Chromosome III
Chromosome IV
Chromosome V
Chromosome VI
Chromosome VII
Chromosome VIII
Chromosome IX
Chromosome X
Chromosome XI
Chromosome XII
Chromosome XIII
Chromosome XIV
Chromosome XV
Chromosome XVI
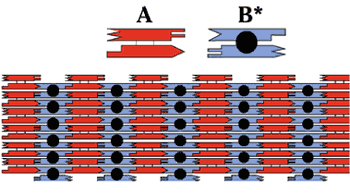
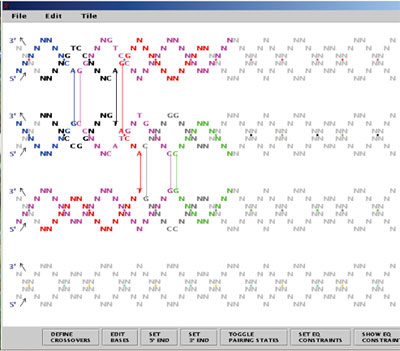
The genetic and physical maps of the sixteen chromosomes of Saccharomyces cerevisiae are based on data presented in Mortimer et al. in preparation (1995) as well as on data presented in earlier reviews (Mortimer and Schild, Microbiol. Rev. 44:519-571 (1980), Microbiol. Rev. 49:181-213 (1985), Mortimer et al., Yeast 5:321-404 (1989), and Mortimer et al., Yeast 8:817-902 (1992). Physical maps were compiled from Riles et al., Genetics 134:81-150 (1992), Riles et al., unpublished data, gene/clone associations reported by many laboratories, and DNA consensus sequences produced by Fred Dietrich from GenBank sequences, and the seven completed yeast chromosomal sequences: I, II, III, V, VIII, IX, and XI (Bussey et al., (1994); Feldmann et al, (1994), EMBL J. in press; Oliver et al. (1992), Nature 357:38-46; Dietrich et al., (1994); Johnston et al., (1994), Science 265:2077-2082 Barrell et al., (1994); Dujon et al. (1994), Nature 369:371-378). All the information in these figures derives directly from the electronic database maintained by theSaccharomyces Genome Database Group at Stanford. Genetic maps are drawn as solid vertical lines; to scale or dashed lines representing linkages established by mitotic crossing over. Centromeres are represented as circles on the genetic map line with the left arm above and the right arm below the centromere. Physical maps are drawn parallel and to the left of the genetic maps Q distinguished by the scale (in kb) to the left of this line. Horizontal tick marks to the right of the genetic map lines indicate the positions of the genes; in crowded regions, these tick marks are joined to the gene symbol by a thin connecting line. Symbols by the physical map represent genes located solely by physical methods. Note that two loci (RDN1 on XII and CUP1 on VIII are known to vary greatly in length from strain to strain. Dashed lines connect the same gene on the genetic and physical maps, only a fraction of such associations are shown. Crowded regions on the genetic map are drawn as expansions. Genes added since Edition 11 are in bold type. Genes that have been mapped on the basis of sequence overlap relative to genetically-mapped genes are positioned to the left of the genetic map line and are connected to this line by tick marks; when the order of these genes relative to the mapped genes is unknown, they are arbitrarily placed distal to the mapped gene. Genes listed below the chromosome maps or expanded regions of these maps have been assigned to the chromosome or region either by genetic or physical methods but have not been positioned more precisely. Some gene symbols in this category are capitalized; this does not necessarily indicate dominance but instead means that the wild type copy was used to map the gene. With this edition the numerous synonyms have not been included. With this edition we have chosen just one symbol for each gene to show on the maps. The symbols all remain in the underlying database. The choice of symbol was guided by an amalgam of consensus, literature usage, clarity relative to function, and priority in the literature. Many of the physical map gene positions are based on hybridization results to the prime clone filter grids. In these cases the location of the gene is arbitarily placed at the center of the hybridizing region. New information on gene mapping or gene name registration should be sent to the Saccharomyces Genome Database Group (FAX: 415-726-7016, electronic mail: yeast-curator@genome.stanford.edu).

http://www.yeastgenome.org/community/mortimer_maps/edition12.shtml
MAPPING CDC MUTATIONS IN THE YEAST S. Cerevisiae
BY RAD52-MEDIATED CHROMOSOME LOSS
Using the chromosome loss-mapping method of Schild and Mortimer, I have
mapped several new temperature-sensitive mutations that define five CDC
genes. Modified procedures were used to facilitate mapping temperature-sen-
sitive mutations in general, and these modifications are discussed. The muta-
tions were assigned to specific chromosomes by chromosome loss procedures,
and linkage relationships were determined subsequently by standard tetrad
analysis. Four of the mutations define new loci. The fifth mutation, cdc63-I,
is shown to be allelic to previously known mutations in the PRTl gene
The results presented here show that the new
procedure of rad52-induced chromosome loss mapping, originally suggested
by D. SCHILD and R. K. MORTIMER (personal communication), is an accurate
and rapid method for assigning mutations to specific chromosomes. Using a
modified rad52-mediated mapping method I have assigned five temperature-
sensitive cdc mutations to specific chromosomes; the results of tetrad analyses
have confirmed the assignments.
The chromosome loss-mapping method used here is based on the finding
that diploids homozygous for rad52 lose chromosomes spontaneously and that
the frequency of loss can be increased by exposure to ionizing radiation (MOR-
TIMER, CONTOPOULOU and SCHILD 1981). Mapping by this method involves
irradiation of rad52/rad52 diploids heterozygous both for the unmapped mu-
tation and for recessive chromosome mutations. Loss of chromosomes results
in the expression of recessive mutations, and correlations observed between
the expression of two mutations can indicate whether the two mutations are
on the same chromosome
http://www.genetics.org/content/110/4/591.full.pdf
Genetic and physical maps of
Saccharomyces cerevisiae
Genetic and physical maps for the 16 chromosomes of Saccharomyces cerevisiae are presented. The genetic map is the result of
40 years of genetic analysis. The physical map was produced from
the results of an international systematic sequencing effort. The
data for the maps are accessible electronically from the Saccharomyces Genome Database (SGD: www.stanford." target="_blank" rel="nofollow">http://genome-www.stanford.
edu/Saccharomyces/).
During the past 40 years, 11 compilations of mapping data for the
yeast Saccharomyces cerevisiae have been made by R. K. Mortimer and colleagues. The last such compilation
included mapping data to 1991, and contained for the first time the results of physical as well as genetic
mapping methods. Here we present the twelfth, and probably the last,
such compilation. These final maps are based on the genetic information
accumulated over the years and, for the physical mapping data, on an
entirely new set of data: the complete genomic sequence of S. cerevisiae.
The genetic and physical maps were derived from two entirely different
types of data. Genetic distances between genes were determined by tetrad
analysis. Distances for gene–gene and gene–centromere linkages are
expressed in centimorgans (cM) and were calculated using a maximumlikelihood equation
which yields values for map distance, an interference parameter, and error calculations for these two parameters. Mapping
results on more than 2,600 named genes are presented. Physical distances
are calculated directly from the complete DNA sequence. The precise values of all parameters (both tetrad analysis results and chromosomal basepair coordinates) are available from the SGD.
Associations between open reading frames (ORFs) and corresponding
mutations were made using a set of hybridization filters, originally produced by L. Riles and M. Olson
13, which are now available from the
American Type Culture Collection (http://www.atcc.org/). Other such
associations were made by complementation experiments using cloned
DNA fragments and/or sequence analysis of mutants. The data for some
of these associations are published, but the documentation for all of them
can be found on SGD.
Now that the entire yeast genome sequence is available, most revisions
of the map will consist of associations between a biological function and
an ORF. These associations will often involve the study of mutants of the
gene. In the past, such an association invariably resulted in the naming of
the gene; this process is likely to continue until all of the genes have been
associated with a function and have thereby acquired a name. Because the
genetic and physical maps are unlikely to change significantly, we see no
need for any future publications; rather, we expect the electronic version of
the maps to evolve into increasingly accurate guides to S. cerevisiae biology.
The maps shown here are also available in a continually updated
electronic form from the SGD (http://genome-www.stanford.edu/
Saccharomyces/), which will also provide directions to other useful
information (gene names, aliases, phenotypes, mapping data, protein
information, and curated compilations of published literature about
genes).
http://www.princeton.edu/genomics/botstein/publications/1997_Cherry_Genetic.pdf
spirits in the material world - the police - ghost in the machine album
There is no political solution
To our troubled evolution
Have no faith in constitution
There is no bloody revolution
We are spirits in the material world
Our so-called leaders speak
With words they try to jail you
They subjugate the meek
But it's the rhetoric of failure
We are spirits in the material world
Where does the answer lie?
Living from day to day
If it's something we can't buy
There must be another way
by I.J. Good who may have known of Yakir's 1964 paper. Of course my first introduction to
back-from-the-future physics was in 1953 as a child in a series of mysterious telephone calls.
President Obama recently gave Yakir a medal for it at The White House - well deserved.
This is an important idea that will change theology as well as physics.
http://www.stardrive.org/
Mysterious phone calls? we'll get back to that but first let me introduce you to Mortimer.
Mortimer is a kind of quite fellow as well. As guides go his function as my "guide" was "financial accountant".
Now this threw me for awhile. I just could not see where money/finance had a place in inter-dimensional guides. Made absolutely no sense to me. Now I thought for a time perhaps this guide would help me with my money and guide me to have abundance. For in fact I was making a pretty good income as a Finance Director. But what in heavens name did that have to do with any of this stuff? What was it that had "value" in a financial sense.
So I looked at the wealthiest and realized most of the truly wealthy retainded one thing they "prized" of value. Money was certainly a way of getting places in a material world. But what was of MORE importance to these "elite" types....you know...the ones who ruled Egypt...the ones who are said to rule kingdoms and basically the world? Seems to me "bloodline" took presidency in monarchies and henceforth were handed down from one member of that line to another eh? Apparently money was the "result" of the thing that had REAL value.... and that was a genetic line. Just as the Merovingian's about bloodline. Or the Queen of England. Ask the elusive "33" what they value and try to get an answer why bloodline/genetics seem so important?
So meet Mortimer my financial accountant....and find out how he balances his books in things of financial "wealth".
"Mortimer" Maps - Edition 12
Chromosome I
Chromosome II
Chromosome III
Chromosome IV
Chromosome V
Chromosome VI
Chromosome VII
Chromosome VIII
Chromosome IX
Chromosome X
Chromosome XI
Chromosome XII
Chromosome XIII
Chromosome XIV
Chromosome XV
Chromosome XVI
The genetic and physical maps of the sixteen chromosomes of Saccharomyces cerevisiae are based on data presented in Mortimer et al. in preparation (1995) as well as on data presented in earlier reviews (Mortimer and Schild, Microbiol. Rev. 44:519-571 (1980), Microbiol. Rev. 49:181-213 (1985), Mortimer et al., Yeast 5:321-404 (1989), and Mortimer et al., Yeast 8:817-902 (1992). Physical maps were compiled from Riles et al., Genetics 134:81-150 (1992), Riles et al., unpublished data, gene/clone associations reported by many laboratories, and DNA consensus sequences produced by Fred Dietrich from GenBank sequences, and the seven completed yeast chromosomal sequences: I, II, III, V, VIII, IX, and XI (Bussey et al., (1994); Feldmann et al, (1994), EMBL J. in press; Oliver et al. (1992), Nature 357:38-46; Dietrich et al., (1994); Johnston et al., (1994), Science 265:2077-2082 Barrell et al., (1994); Dujon et al. (1994), Nature 369:371-378). All the information in these figures derives directly from the electronic database maintained by theSaccharomyces Genome Database Group at Stanford. Genetic maps are drawn as solid vertical lines; to scale or dashed lines representing linkages established by mitotic crossing over. Centromeres are represented as circles on the genetic map line with the left arm above and the right arm below the centromere. Physical maps are drawn parallel and to the left of the genetic maps Q distinguished by the scale (in kb) to the left of this line. Horizontal tick marks to the right of the genetic map lines indicate the positions of the genes; in crowded regions, these tick marks are joined to the gene symbol by a thin connecting line. Symbols by the physical map represent genes located solely by physical methods. Note that two loci (RDN1 on XII and CUP1 on VIII are known to vary greatly in length from strain to strain. Dashed lines connect the same gene on the genetic and physical maps, only a fraction of such associations are shown. Crowded regions on the genetic map are drawn as expansions. Genes added since Edition 11 are in bold type. Genes that have been mapped on the basis of sequence overlap relative to genetically-mapped genes are positioned to the left of the genetic map line and are connected to this line by tick marks; when the order of these genes relative to the mapped genes is unknown, they are arbitrarily placed distal to the mapped gene. Genes listed below the chromosome maps or expanded regions of these maps have been assigned to the chromosome or region either by genetic or physical methods but have not been positioned more precisely. Some gene symbols in this category are capitalized; this does not necessarily indicate dominance but instead means that the wild type copy was used to map the gene. With this edition the numerous synonyms have not been included. With this edition we have chosen just one symbol for each gene to show on the maps. The symbols all remain in the underlying database. The choice of symbol was guided by an amalgam of consensus, literature usage, clarity relative to function, and priority in the literature. Many of the physical map gene positions are based on hybridization results to the prime clone filter grids. In these cases the location of the gene is arbitarily placed at the center of the hybridizing region. New information on gene mapping or gene name registration should be sent to the Saccharomyces Genome Database Group (FAX: 415-726-7016, electronic mail: yeast-curator@genome.stanford.edu).
http://www.yeastgenome.org/community/mortimer_maps/edition12.shtml
MAPPING CDC MUTATIONS IN THE YEAST S. Cerevisiae
BY RAD52-MEDIATED CHROMOSOME LOSS
Using the chromosome loss-mapping method of Schild and Mortimer, I have
mapped several new temperature-sensitive mutations that define five CDC
genes. Modified procedures were used to facilitate mapping temperature-sen-
sitive mutations in general, and these modifications are discussed. The muta-
tions were assigned to specific chromosomes by chromosome loss procedures,
and linkage relationships were determined subsequently by standard tetrad
analysis. Four of the mutations define new loci. The fifth mutation, cdc63-I,
is shown to be allelic to previously known mutations in the PRTl gene
The results presented here show that the new
procedure of rad52-induced chromosome loss mapping, originally suggested
by D. SCHILD and R. K. MORTIMER (personal communication), is an accurate
and rapid method for assigning mutations to specific chromosomes. Using a
modified rad52-mediated mapping method I have assigned five temperature-
sensitive cdc mutations to specific chromosomes; the results of tetrad analyses
have confirmed the assignments.
The chromosome loss-mapping method used here is based on the finding
that diploids homozygous for rad52 lose chromosomes spontaneously and that
the frequency of loss can be increased by exposure to ionizing radiation (MOR-
TIMER, CONTOPOULOU and SCHILD 1981). Mapping by this method involves
irradiation of rad52/rad52 diploids heterozygous both for the unmapped mu-
tation and for recessive chromosome mutations. Loss of chromosomes results
in the expression of recessive mutations, and correlations observed between
the expression of two mutations can indicate whether the two mutations are
on the same chromosome
http://www.genetics.org/content/110/4/591.full.pdf
Genetic and physical maps of
Saccharomyces cerevisiae
Genetic and physical maps for the 16 chromosomes of Saccharomyces cerevisiae are presented. The genetic map is the result of
40 years of genetic analysis. The physical map was produced from
the results of an international systematic sequencing effort. The
data for the maps are accessible electronically from the Saccharomyces Genome Database (SGD: www.stanford." target="_blank" rel="nofollow">http://genome-www.stanford.
edu/Saccharomyces/).
During the past 40 years, 11 compilations of mapping data for the
yeast Saccharomyces cerevisiae have been made by R. K. Mortimer and colleagues. The last such compilation
included mapping data to 1991, and contained for the first time the results of physical as well as genetic
mapping methods. Here we present the twelfth, and probably the last,
such compilation. These final maps are based on the genetic information
accumulated over the years and, for the physical mapping data, on an
entirely new set of data: the complete genomic sequence of S. cerevisiae.
The genetic and physical maps were derived from two entirely different
types of data. Genetic distances between genes were determined by tetrad
analysis. Distances for gene–gene and gene–centromere linkages are
expressed in centimorgans (cM) and were calculated using a maximumlikelihood equation
which yields values for map distance, an interference parameter, and error calculations for these two parameters. Mapping
results on more than 2,600 named genes are presented. Physical distances
are calculated directly from the complete DNA sequence. The precise values of all parameters (both tetrad analysis results and chromosomal basepair coordinates) are available from the SGD.
Associations between open reading frames (ORFs) and corresponding
mutations were made using a set of hybridization filters, originally produced by L. Riles and M. Olson
13, which are now available from the
American Type Culture Collection (http://www.atcc.org/). Other such
associations were made by complementation experiments using cloned
DNA fragments and/or sequence analysis of mutants. The data for some
of these associations are published, but the documentation for all of them
can be found on SGD.
Now that the entire yeast genome sequence is available, most revisions
of the map will consist of associations between a biological function and
an ORF. These associations will often involve the study of mutants of the
gene. In the past, such an association invariably resulted in the naming of
the gene; this process is likely to continue until all of the genes have been
associated with a function and have thereby acquired a name. Because the
genetic and physical maps are unlikely to change significantly, we see no
need for any future publications; rather, we expect the electronic version of
the maps to evolve into increasingly accurate guides to S. cerevisiae biology.
The maps shown here are also available in a continually updated
electronic form from the SGD (http://genome-www.stanford.edu/
Saccharomyces/), which will also provide directions to other useful
information (gene names, aliases, phenotypes, mapping data, protein
information, and curated compilations of published literature about
genes).
http://www.princeton.edu/genomics/botstein/publications/1997_Cherry_Genetic.pdf


















 \
\
















 The Trustees of the British Museum
The Trustees of the British Museum



 .
.